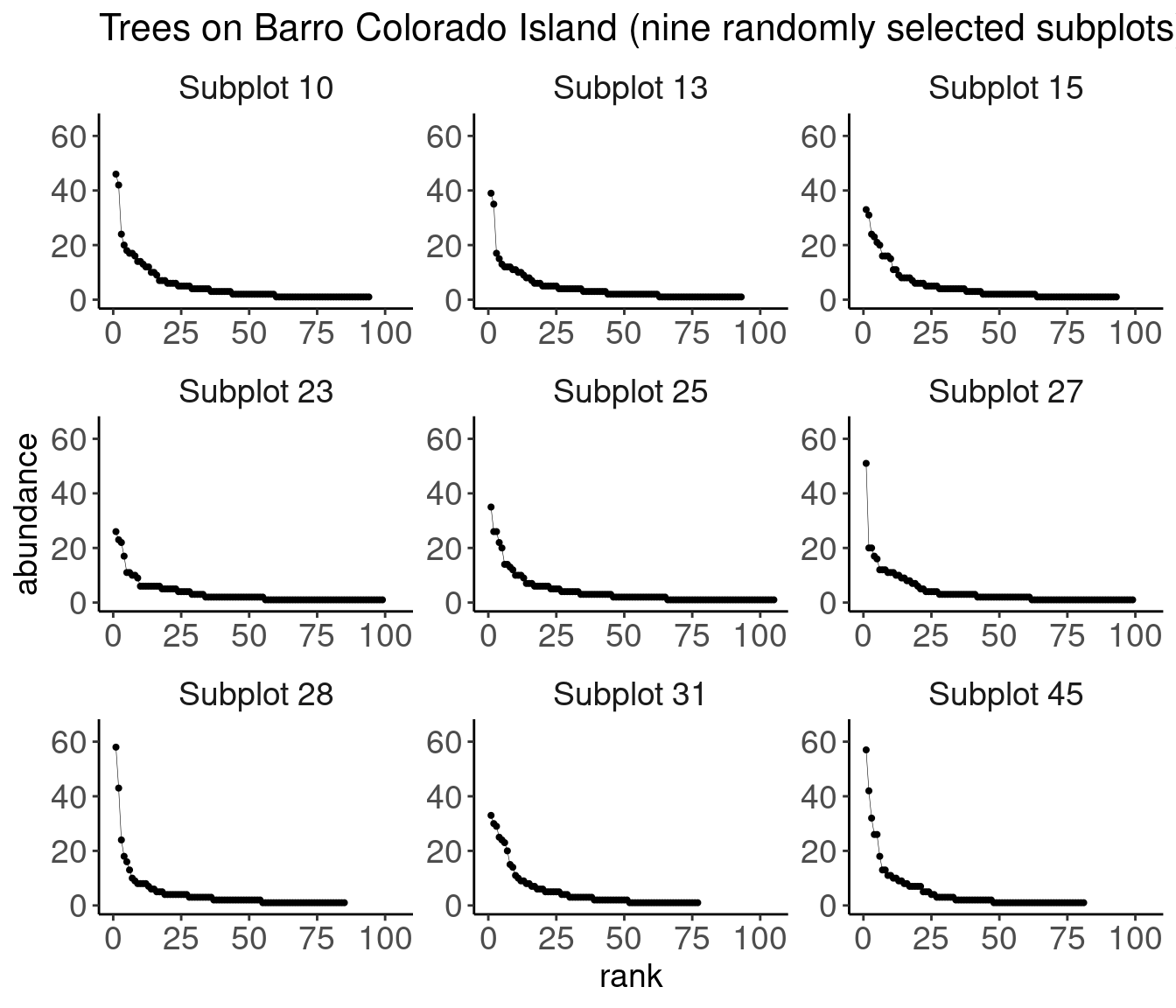
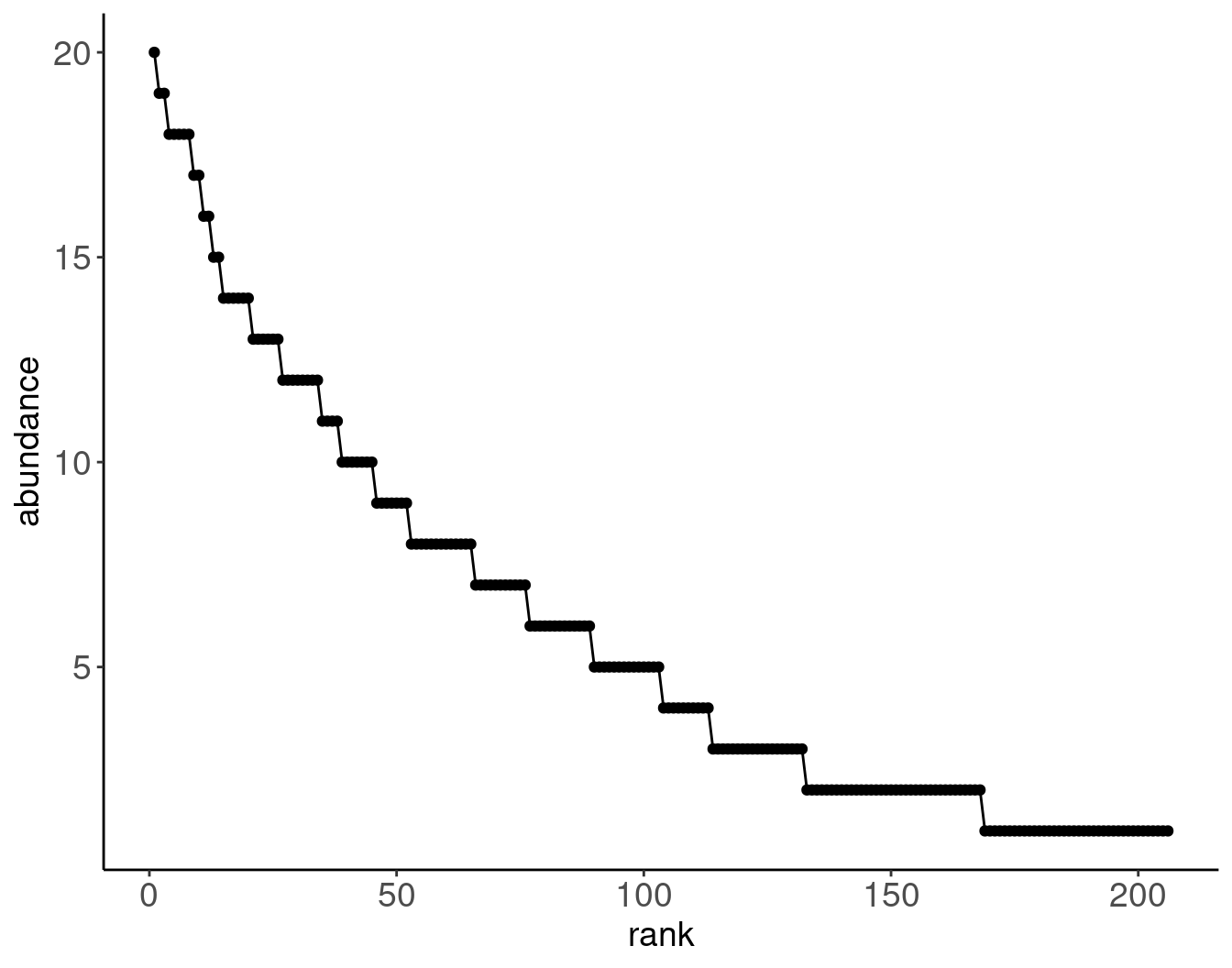
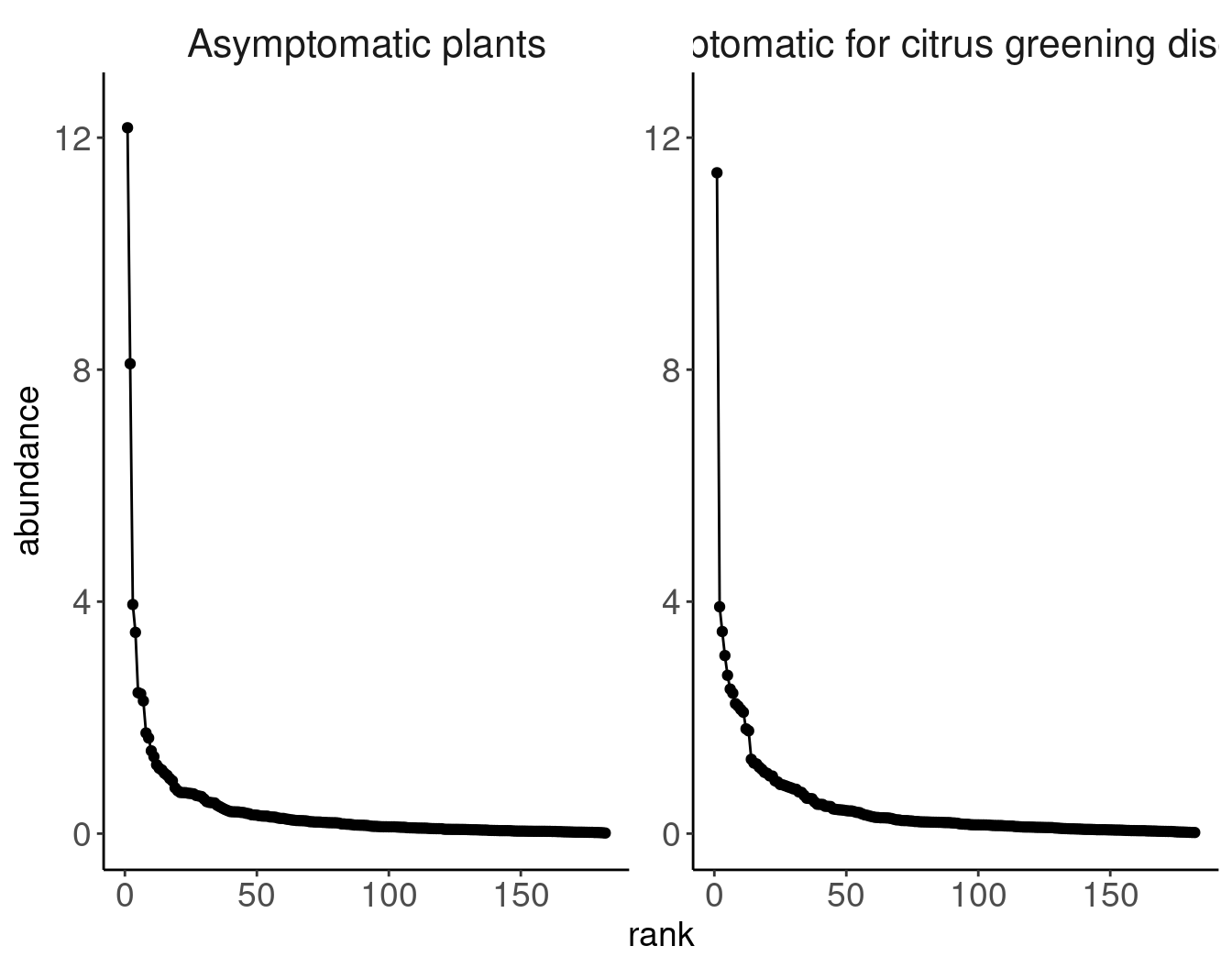
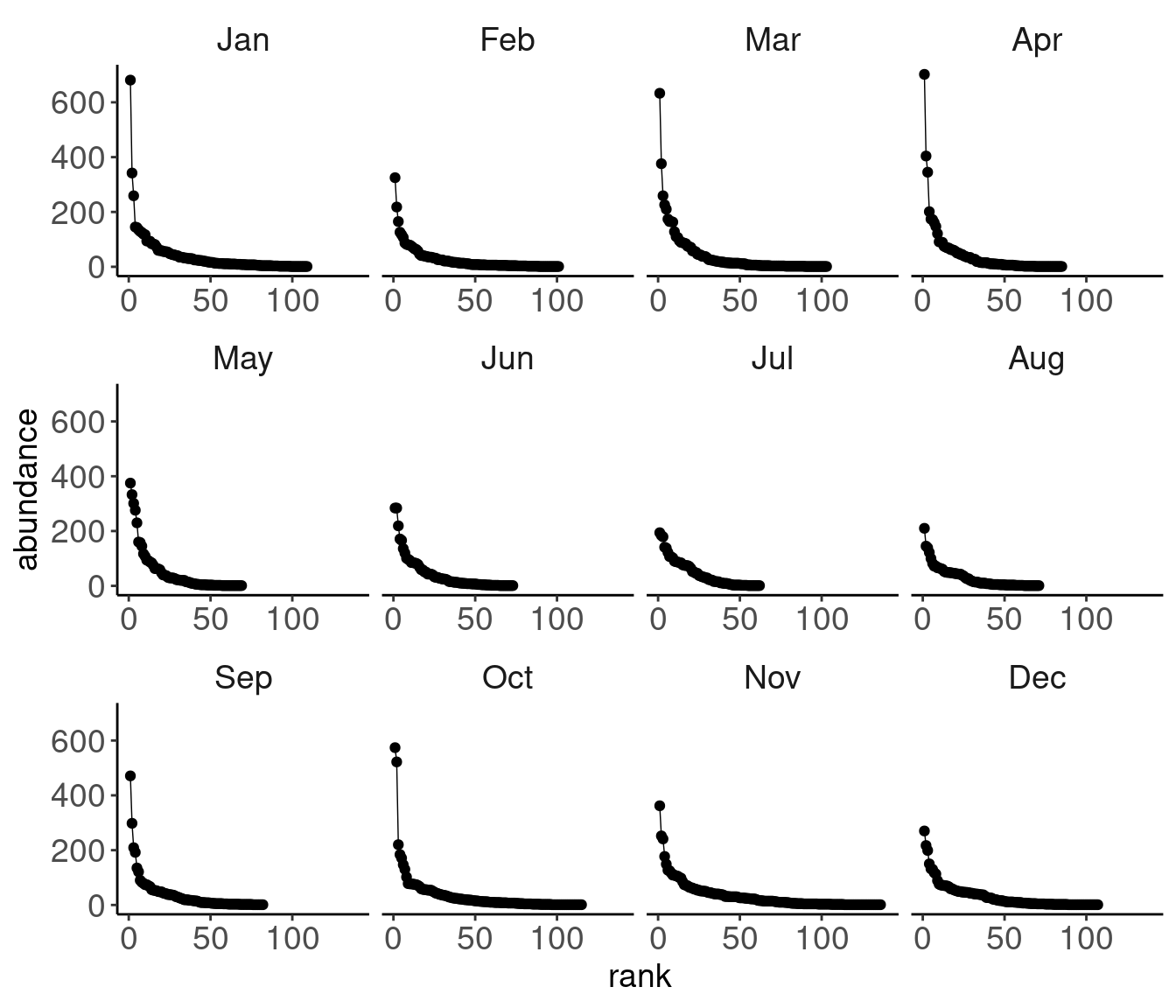
Example 2: Lepidoptera at a light trap in Maine, USA.
Code
# Data transcribed from Bonson 1964, 'Patterns in the balance of nature and related problems in quantitative ecology'", accessed through archive.orgtribble(~abundance, ~nsp,1,38,2,36,3,19,4,10,5,14,6,13,7,11,8,13,9,7,10,7,11,4,12,8,13,6,14,6,15,2,16,2,17,2,18,5,19,2,20,1) |>uncount(nsp) |>mutate(spid =row_number()) |>mutate(rank =rank(desc(abundance), ties.method ="first")) |>ggplot(aes(x = rank, y = abundance)) +geom_point() +geom_line()
Example 3: Bacteria in citrus roots
Code
# Data downloaded from supplement of # Blaustein et al. "Defining the Core Citrus Leaf- and Root-Associated Microbiota: Factors Associated with Community Structure and Implications for Managing Huanglongbing (Citrus Greening) Disease",# https://doi.org/10.1128/AEM.00210-17# First 46 rows have data from phyllosphere (leaf) bacteriaread_excel("hlb-microbes.xlsx", skip =47) |> dplyr::rename(meanabun_asym =11, meanabun_sym =13) |>select(starts_with("meanabun_")) |>pivot_longer(1:2, values_to ="abundance") |>mutate(name =ifelse(name =="meanabun_asym", "Asymptomatic plants","Symptomatic for citrus greening disease")) |>group_by(name) |>mutate(rank =rank(desc(abundance), ties.method ="first")) |>ggplot(aes(x = rank, y = abundance)) +geom_point() +geom_line() +facet_wrap(.~name, scales ="free") +ylim(0,12.5) +theme(strip.text =element_text(size =16))
Example 4: Birds recorded at GKVK campus in 2024
Code
# Uncomment the following lines to download data from ebird, # and add your ebird key# library(rebird)# gkvk_2024 <- tibble(date_format = na.omit(ymd(paste0("2024-",rep(1:12, each = 31),"-",01:31))),# ebird_out = map(date_format,# ~ebirdhistorical(loc = 'L2525709', # for GKVK campus# date = .x,# key = "<your ebird key>")))gkvk_2024 <-readRDS("gkvk_birds.rds")list_rbind(gkvk_2024$ebird_out) |>mutate(month =month(ymd_hm(obsDt), label = T, abbr = T)) |>group_by(comName,sciName,locName, month) |>summarize(abundance =sum(howMany)) |>ungroup() |>group_by(month) |>mutate(rank =rank(desc(abundance), ties.method ="first")) |>ggplot(aes(x = rank, y = abundance)) +geom_point(aes(group = comName)) +geom_line(linewidth =0.25) +facet_wrap(.~month, scales ="free_x") +xlim(0,140)## `summarise()` has grouped output by 'comName', 'sciName', 'locName'. You can## override using the `.groups` argument.## Warning: Removed 42 rows containing missing values or values outside the scale range## (`geom_point()`).
Think and discuss:
What might cause a species to be common or rare in a community?
What process(es) might give rise to this community-level pattern of a “long tail” in the distribution?
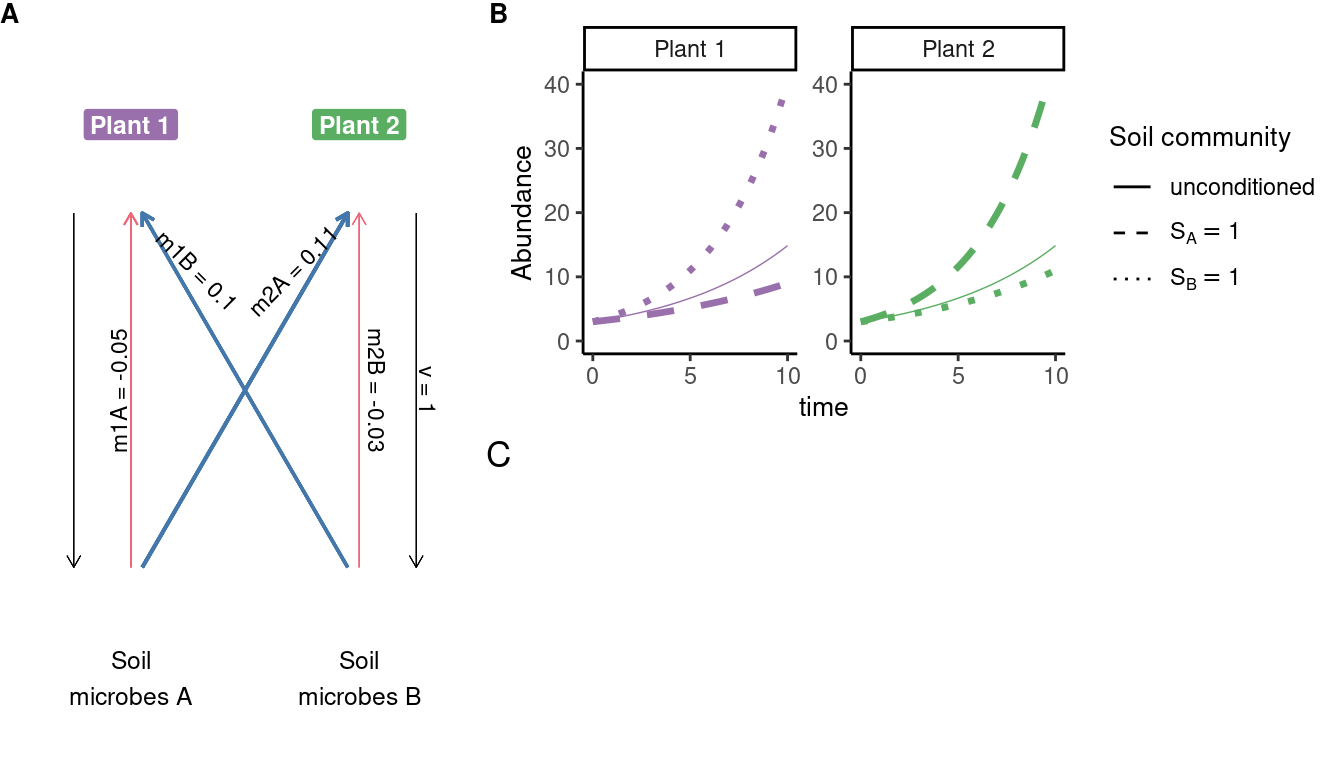
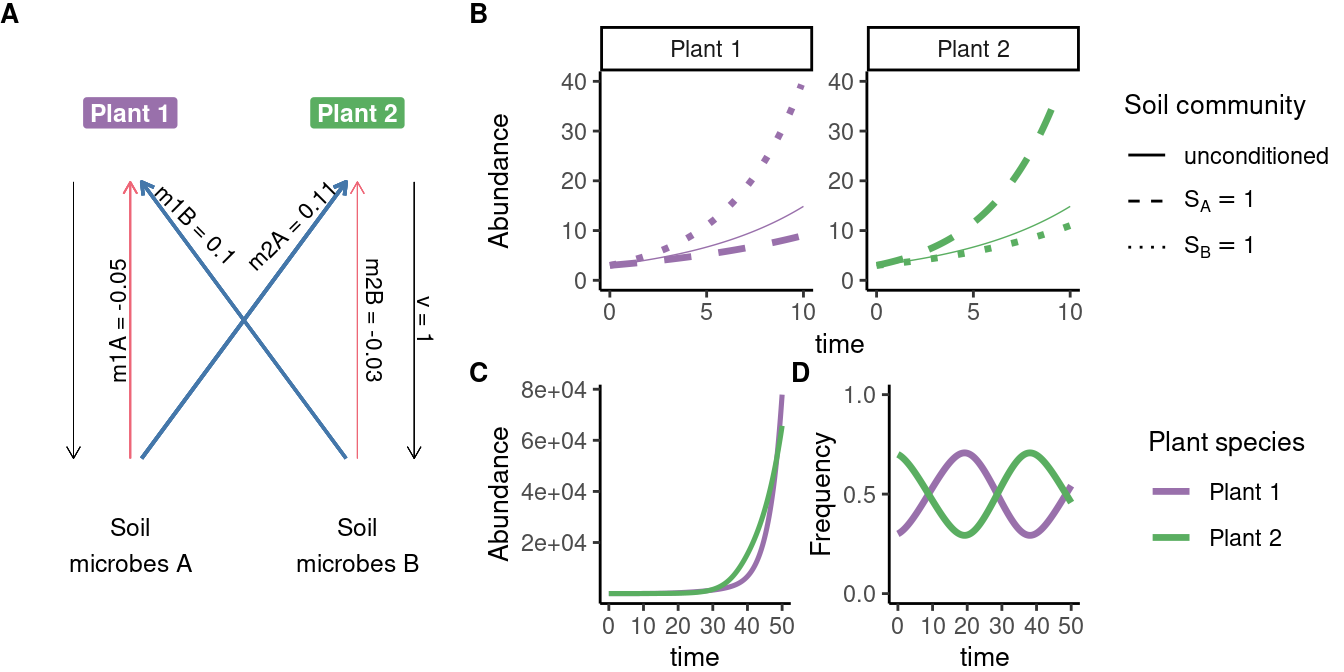
Interpretation: Soil communities grow at a rate determined by the frequency of each plant (and the relative strength of each plant’s conditioning effect, \(\nu\)).
Overview of model dynamics
Plant population growth rates depend on the composition of the microbial community
Microbial community composition in turn depends on the relative frequency of each plant.
We can express the system’s dynamics in terms of plant and soil frequencies.
source("simulate-psf-model.R")## Loading required package: deSolve## Loading required package: rootSolve## Loading required package: latex2exp## ## Attaching package: 'latex2exp'## The following object is masked from 'package:plotly':## ## TeX## Warning: The `legend.text.align` argument of `theme()` is deprecated as of ggplot2## 3.5.0.## ℹ Please use theme(legend.text = element_text(hjust)) instead.fig1_p1 <- fig1$p5fig1_p1[[2]][[2]] =NULLfig1_p1 ## Warning in geom_segment(aes(x = 0, xend = 0, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.05, xend = 0.95, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.95, xend = 0.05, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1, xend = 1, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = -0.25, xend = -0.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1.25, xend = 1.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning: Removed 6 rows containing missing values or values outside the scale range## (`geom_line()`).
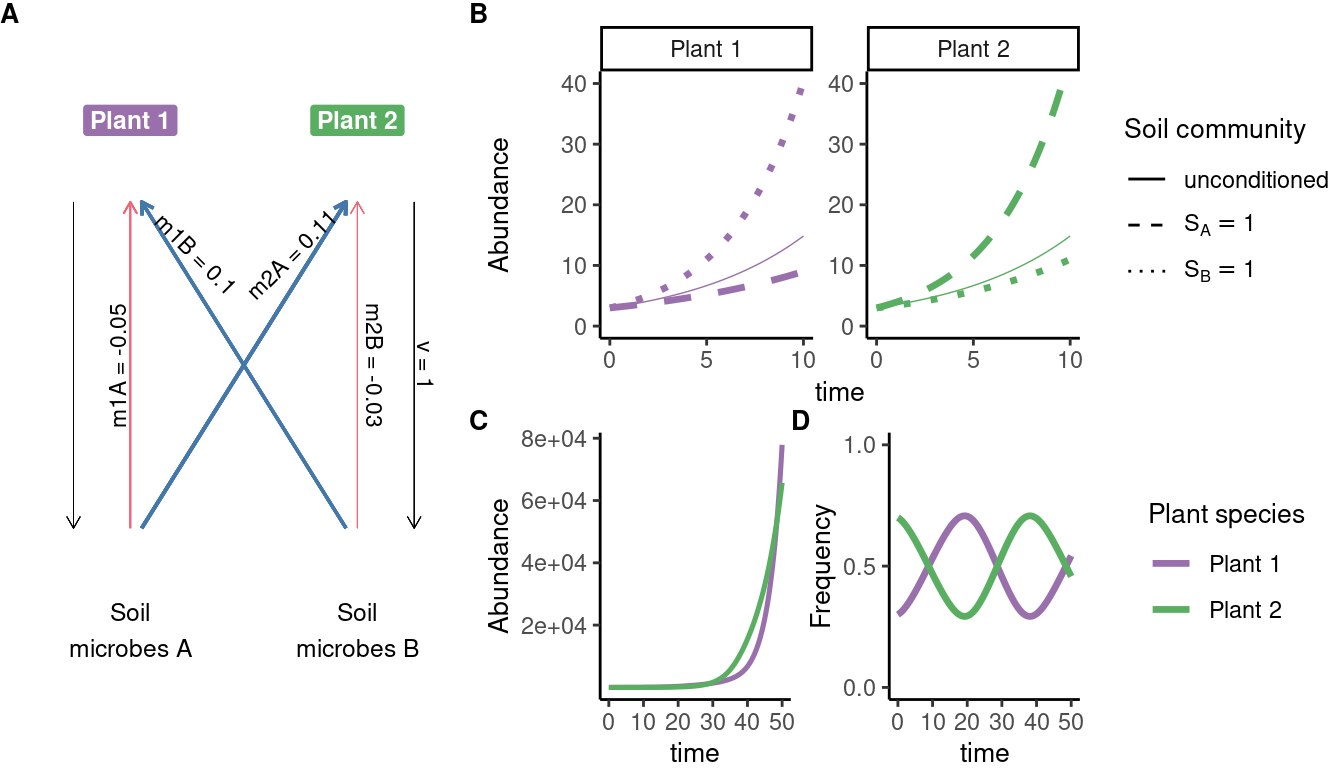
Visualization of model dynamics
Code
fig1$p5## Warning in geom_segment(aes(x = 0, xend = 0, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.05, xend = 0.95, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.95, xend = 0.05, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1, xend = 1, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = -0.25, xend = -0.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1.25, xend = 1.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning: Removed 6 rows containing missing values or values outside the scale range## (`geom_line()`).
Under this set of parameters, the frequencies of the species oscillate - a form of neutral coexistence.
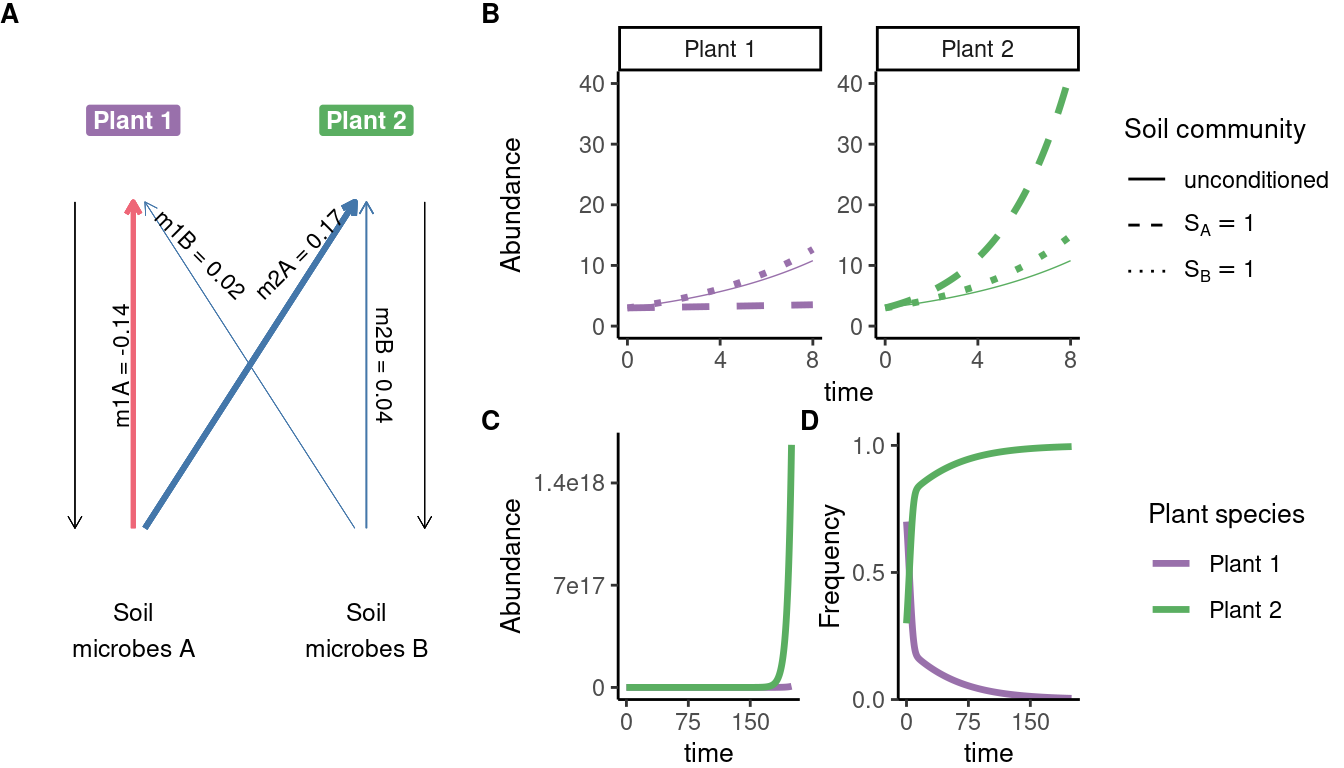
Visualization of model dynamics
Code
fig2AtoD## Warning in geom_segment(aes(x = 0, xend = 0, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.05, xend = 0.95, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.95, xend = 0.05, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1, xend = 1, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = -0.25, xend = -0.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1.25, xend = 1.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning: Removed 122 rows containing missing values or values outside the scale range## (`geom_line()`).
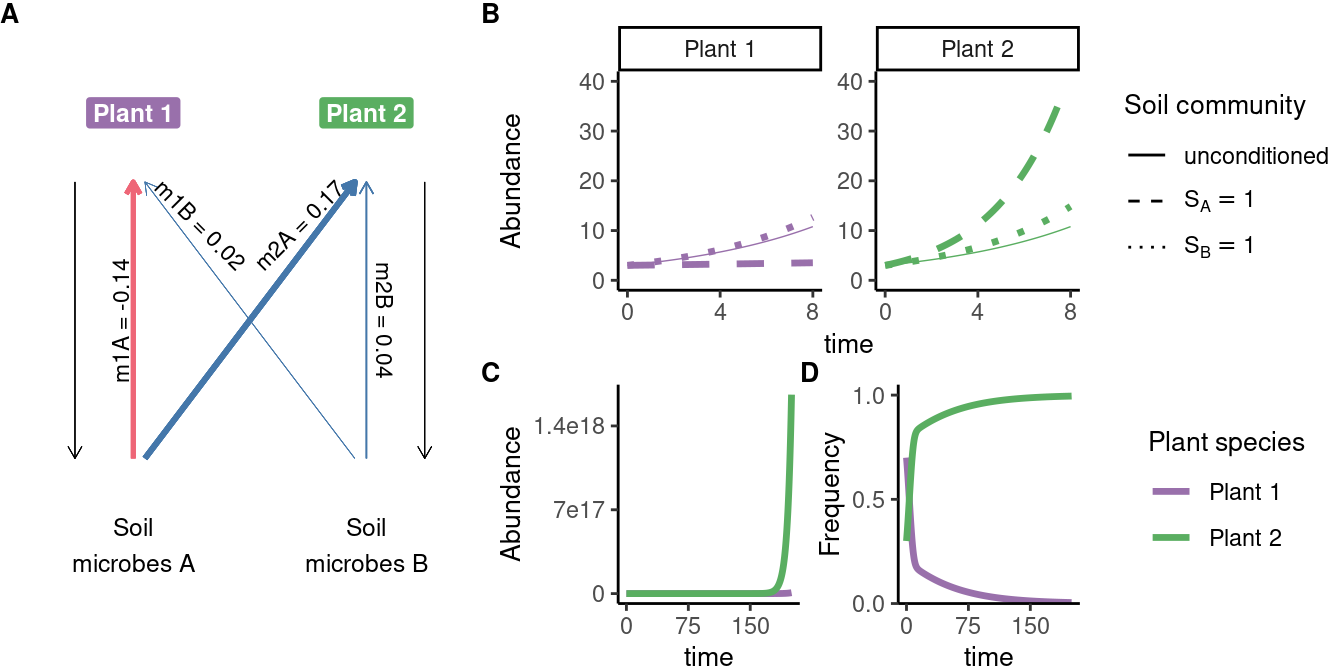
Under other parameters, the frequency of one species reaches 1 - a form of species exclusion.
What determines the outcome of these interactions?
Original analysis emphasized the role of species–specific pathogens (or mutualists that favor the non-host species)
What determines the outcome of these interactions?
Original analysis emphasized the role of species–specific pathogens (or mutualists that favor the non-host species)
In what ways is this model similar to or different from the Lotka-Volterra framework?
What biological processes are included in the model, and what are missing?
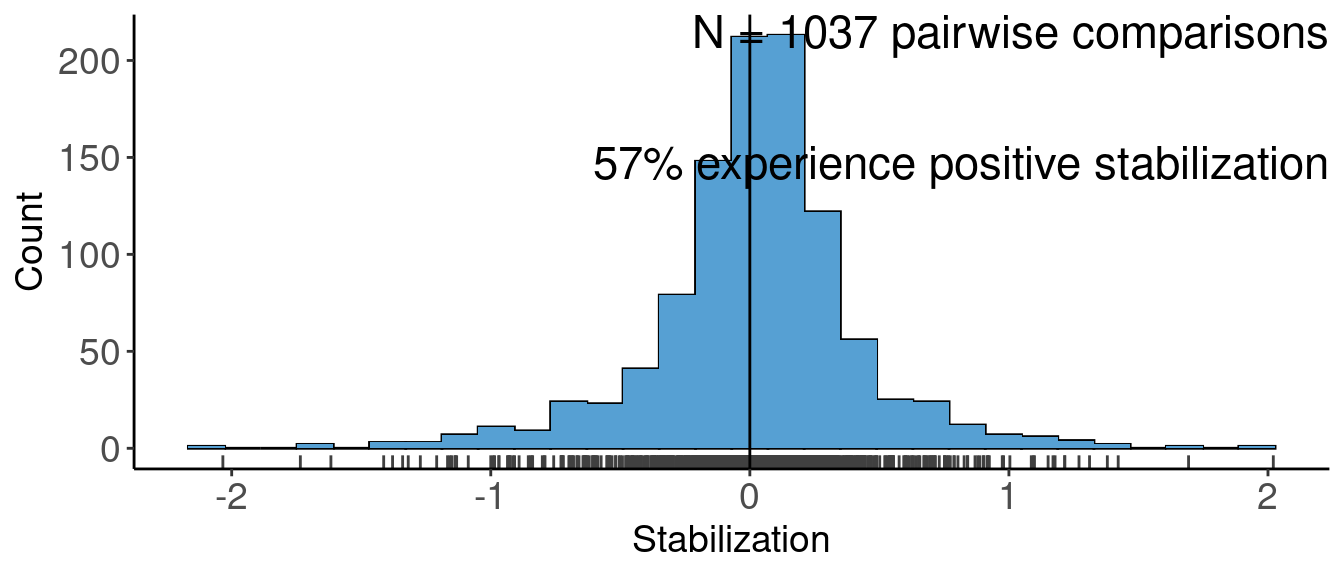
Lasting influence of the Bever model
Code
# Dataset downloaded from Figshare: # curl::curl_download("https://figshare.com/ndownloader/files/14874749", destfile = "crawford-supplement.xlsx")read_excel("crawford-supplement.xlsx", sheet ="Data") |>mutate(stab =-0.5*rrIs) |># show in terms of stabilization instead of I_sfilter(stab >-3) |>ggplot(aes(x = stab)) +geom_histogram(color ="black") +geom_histogram(fill ="#56A0D3") +geom_rug(color ="grey25") +geom_vline(xintercept =0) +annotate("text", x =Inf, y =Inf, label ="N = 1037 pairwise comparisons\n 57% experience positive stabilization",vjust=1, hjust =1, size =6) +xlab("Stabilization") +ylab("Count")## `stat_bin()` using `bins = 30`. Pick better value with `binwidth`.## `stat_bin()` using `bins = 30`. Pick better value with `binwidth`.
How does this relate to species commonness and rarity?
“In 1997, a big change happened in my approach to studying plant–microbe interactions. While attending the Mycological Society of America meeting in Montreal, I listened to Jim Bever talk about a simple but powerful framework on plant–soil feedback. He showed how his concept could be used to study plant and microbial effects and responses at the same time.”
# data extracted from Fig. 3 of Klironomos 2002, https://www.nature.com/articles/417067a.pdf# using WebPlotDigitizertribble(~fb, ~abun,-0.34, 3.27,-0.41, 7.94,-0.27, 2.20,0.01, 41.49,-0.14, 27.39,-0.05, 67.29,0.00, 59.28,-0.16, 7.81,0.11, 35.77,-0.08, 42.83,-0.15, 1.63,-0.36, 1.22,-0.07, 1.33,-0.27, 5.30,-0.40, 2.27,0.06, 87.34,-0.21, 4.75,-0.23, 0.38,-0.02, 35.58,-0.05, 41.78,-0.25, 1.67,-0.20, 7.32,-0.15, 12.71,-0.09, 10.36,-0.18, 11.18,0.04, 53.85,-0.29, 6.85,-0.14, 31.00,-0.33, 3.01,-0.07, 15.24,-0.15, 67.60,0.00, 14.95,-0.22, 28.98,0.11, 50.46,-0.03, 51.82,-0.23, 18.16,-0.15, 0.34,-0.36, 0.44,-0.30, 1.19,0.20, 62.78,-0.30, 7.89,-0.14, 21.72,-0.37, 5.60,-0.13, 64.76,-0.26, 0.91,-0.16, 45.70,-0.24, 6.83,-0.43, 2.80,-0.13, 29.19,-0.49, 0.77,-0.05, 13.94,-0.06, 37.92,0.01, 68.30,-0.36, 10.76,-0.31, 7.38,-0.27, 4.26,0.06, 60.02,0.04, 71.89,-0.23, 0.63,-0.01, 80.93,) |>ggplot(aes(x = fb, y = abun)) +geom_point(size =2.5) +# fit line from Fig 3 of the papergeom_function(fun =function(x) 114.529*x^2+156.652*x +44.013) +xlab("Species-level feedback") +ylab("Relative abundance")
Species–level feedback calculated as (approx.)
feedbacki = biomassi in soil i - biomassi in other soil
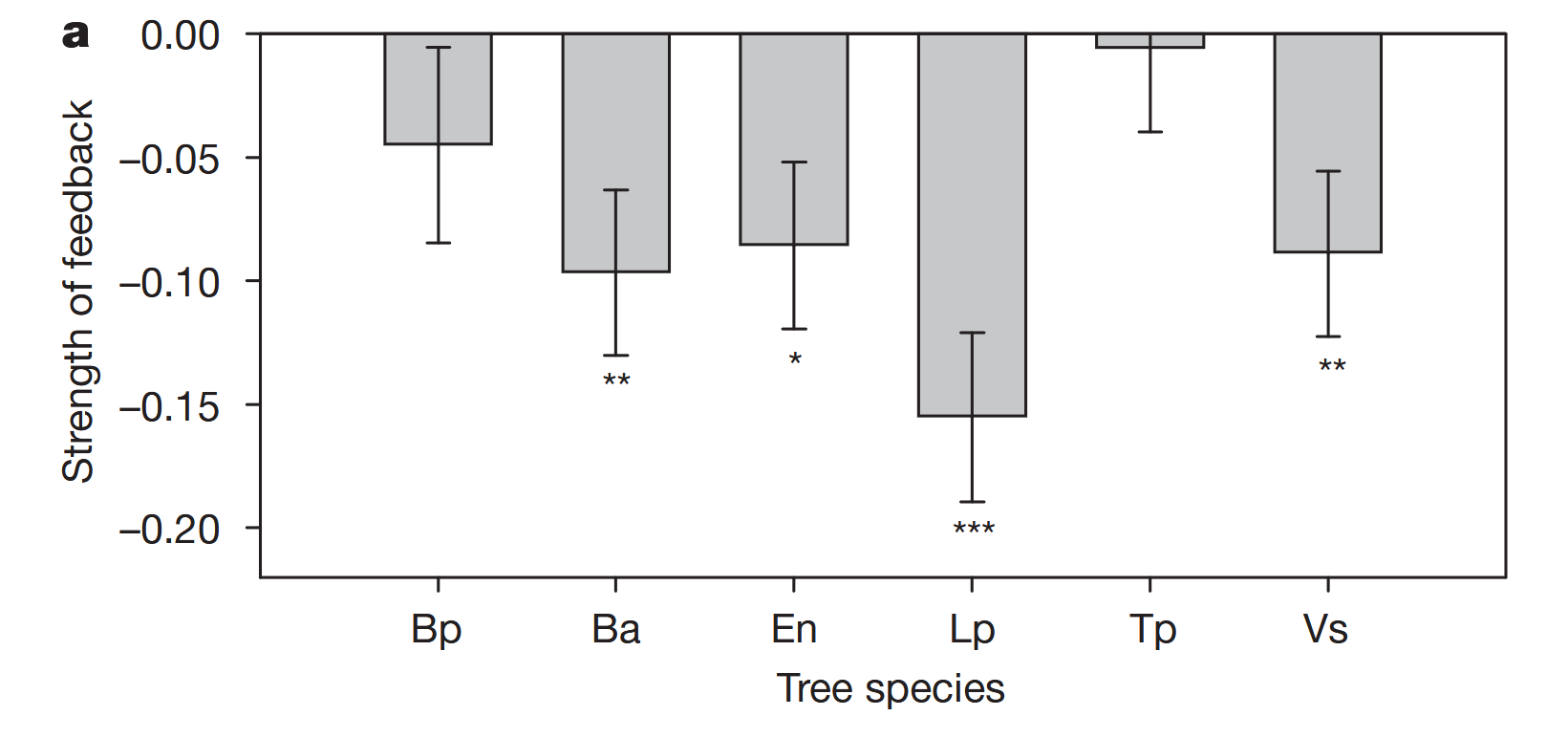
Approach: Shade-house studies of pairwise feedback among 6 species of varying abundances, and a complementary field transplant experiment. (Shade-house study with soils from BCI; field study in another nearby forest).
Experimentally quantified pairwise feedback (\(I_S\)) between \({6 \choose 2} = 15\) pairs of species.
For each focal species, calculate the average of all \(I_S\) involving that species:
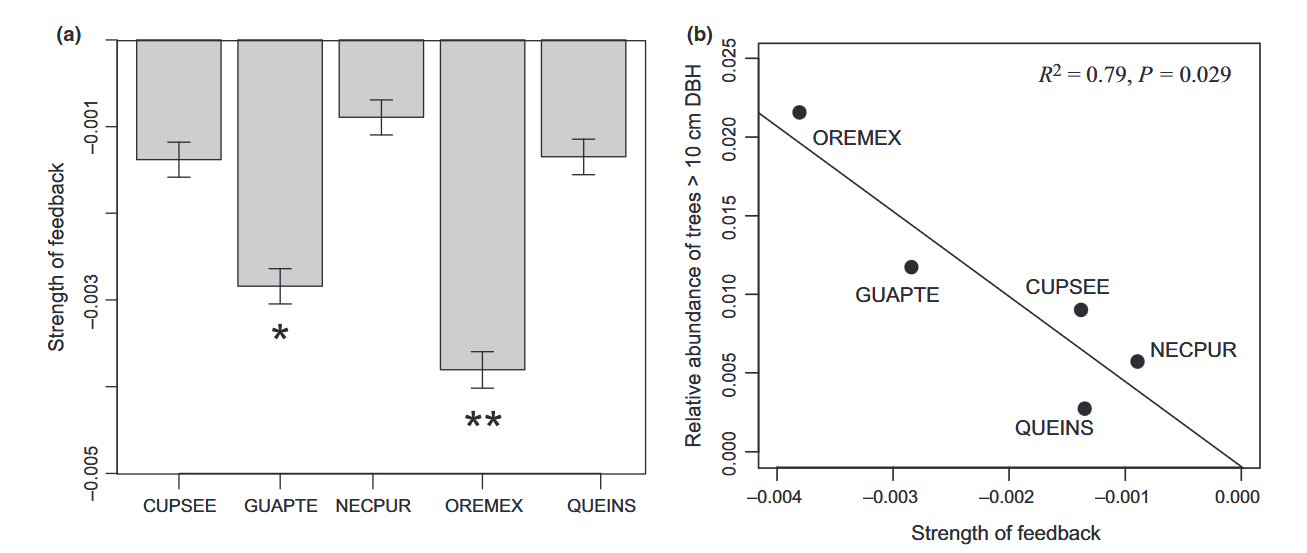
Key result: Species involved in more stabilized interactions (more negative \(I_S\)) tend to be less abundant in the forest
Complementary field study provided further support
“Our [studies] reinforce the conclusion that trees are abundant because they are less susceptible to the detrimental effects of their associated soil communities than are rarer tree species. Thus, localized negative plant–soil feedback occurring between plants and below-ground organisms may be a general mechanism for the maintenance of plant species diversity and patterns of relative abundance across ecosystems ranging from temperate grasslands to tropical forests.”
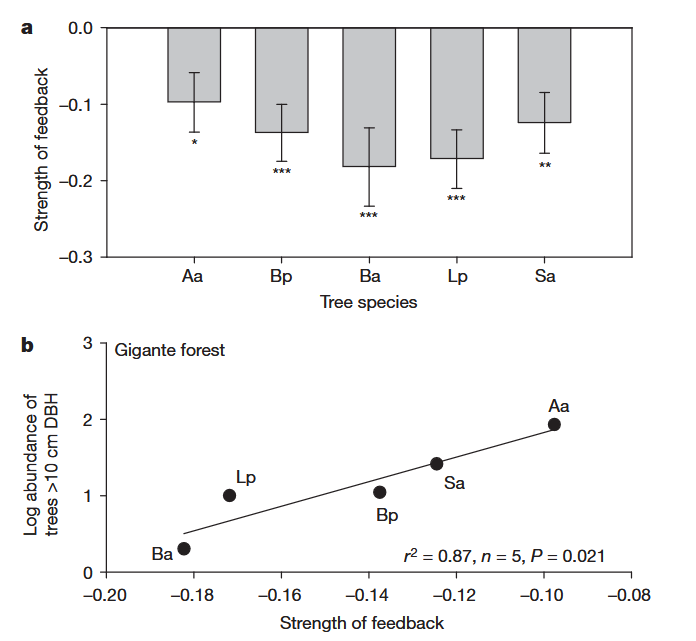
Approach: Statistical analysis of seedling census data from BCI to evlauate the factors that determine seedling survival.
Key result: Species that have stronger self-limitation in seedling recruitment are less common as adults in the forest.
Understanding the drivers of species abundance is critical for identifying and protecting rare species that have an inherently higher risk of extinction. Previous effort to identify key traits that correlate with species rarity have had limited success. The results here suggest that such studies should look beyond morphological and physiological traits to include species sensitivity to biotic interactions, namely, the degree to which species inhibit their own regeneration.
Let’s reflect:
What is the conceptual argument built up through these studies?
Do you think this is a strong mechanistic explanation?
What more evidence would you like to see to further support or reject this hypothesis?
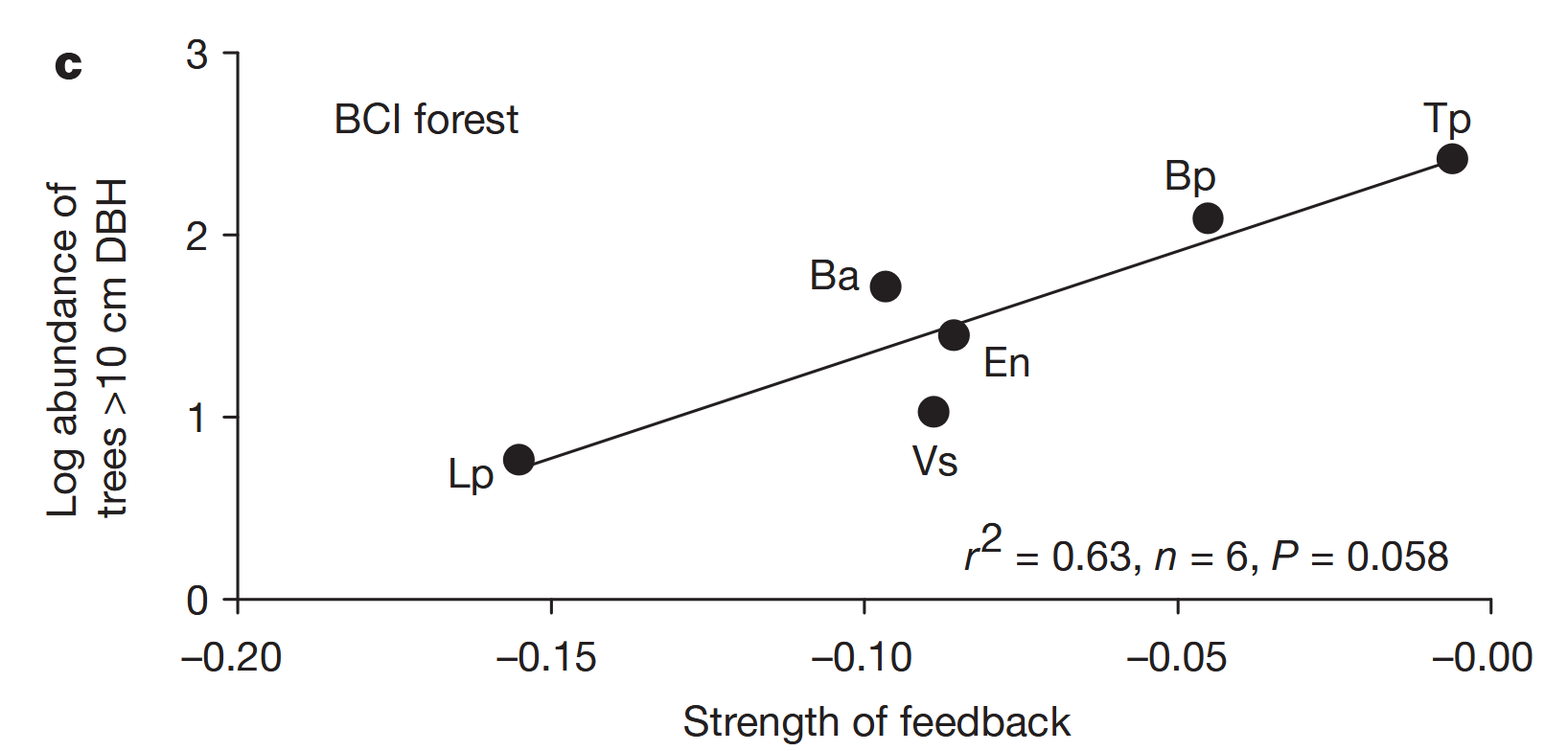
A wrinkle in the story
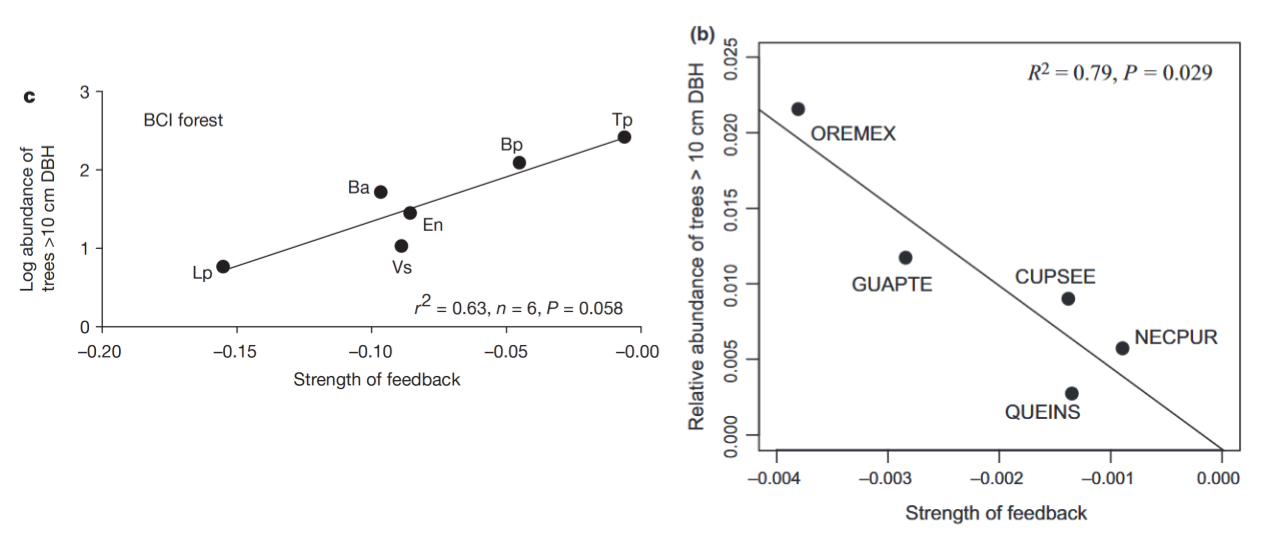
Approach: Shade-house study of pairwise feedback among 5 species of varying abundances, along with extensive field evaluation of mycorrhizal associations. Work done in lower montane tropical forests in Panama.
Key result: Species involved in more stabilized interactions (more negative \(I_S\)) tend to be more abundant in forests
Contrasting relationships across two nearby forests
“This result contrasts with findings from lowland tropical forest and temperate grasslands, where more abundant species show weaker negative PSF. This is the first PSF experiment ever done including montane tree species and more research in this topic is needed to confirm if this is widespread phenomenon of just a peculiarity of our system.”
Summary of timeline
1997: Bever’s feedback model, which suggested an experiment design and metric of stabilization
2002: Klironomos’ study, correlating microbial responses to abundance in a temperate old field
2010: Mangan et al.’s study, correlating strength of pairwise feedback to abundance in a tropical forest
2010: Comita et al.’s study, correlating strength of neighborhood effect to abundance in the same tropical forest
2016: Corrales et al.’s study, finding the opposite correlation between strength of pairwise feedback and abundance in a tropical montane forest
Let’s reflect:
What do you think might be happening here?
Theoretical advances in feedback theory
Recall that the original analysis in 1997 emphasized the strength of intra- vs. interspecific interactions:
\(I_S\) does not fully capture microbial effects, even in this simple model
With equivalent values of \(I_S\), vastly different model dynamics are possible.
Code
fig1$p5## Warning in geom_segment(aes(x = 0, xend = 0, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.05, xend = 0.95, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.95, xend = 0.05, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1, xend = 1, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = -0.25, xend = -0.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1.25, xend = 1.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning: Removed 6 rows containing missing values or values outside the scale range## (`geom_line()`).
With equivalent values of \(I_S\), vastly different model dynamics are possible.
Code
fig2AtoD## Warning in geom_segment(aes(x = 0, xend = 0, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.05, xend = 0.95, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 0.95, xend = 0.05, y = 0.1, yend = 0.9), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1, xend = 1, y = 0.1, yend = 0.9), arrow = arrow(length = unit(0.03, : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = -0.25, xend = -0.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in geom_segment(aes(x = 1.25, xend = 1.25, y = 0.9, yend = 0.1), : All aesthetics have length 1, but the data has 4 rows.## ℹ Please consider using `annotate()` or provide this layer with data containing## a single row.## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning in is.na(x): is.na() applied to non-(list or vector) of type## 'expression'## Warning: Removed 122 rows containing missing values or values outside the scale range## (`geom_line()`).
Think back to how \(I_S\) was derived - what does it guarantee?
\(I_S<0\) suggests neutral stability (in frequency space)
Species coexistence if \(\frac{dF_1}{dt}|_{F_2 = 1} > 0\) and \(\frac{dF_2}{dt}|_{F_1 = 1} > 0\)
Independent exercise: Derive the criteria for coexistence in the Bever 1997 model through analyzing feasibility analysis and through invasion analysis.
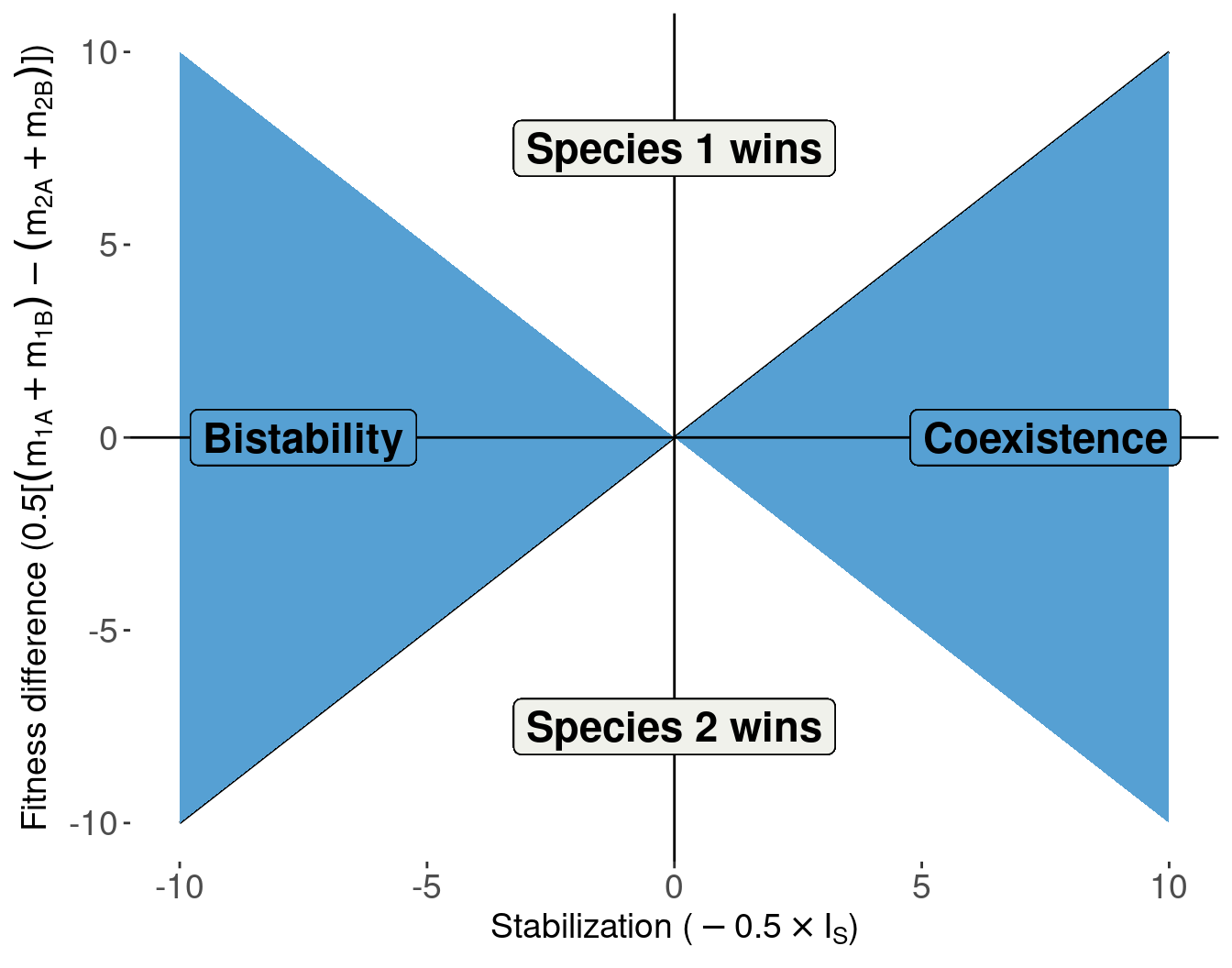
We can also express this in the “Chesson” language
Decompose the invasion growth rates into the strength of stabilization and fitness differences.
Empirically, microbially-mediated fitness differences tend to outweight stabilizing effects
(Data shown for 72 out of >500 species pairs in the meta-analysis)
Open question:
Is variation in relative abundance better explained by variation in species’ overall responses to microbes (fitness imbalance) than to “self vs. other” microbes (stabilization)?
The Bever 1997 framework does not permit multispecies coexistence
Here, we extend this canonical model of PSFs to include an arbitrary number of plant species and analyse the dynamics. Surprisingly, we find that coexistence of more than two species is virtually impossible, suggesting that alternative theoretical frameworks are needed to describe feedbacks observed in diverse natural communities.
The [abundance] projected onto [frequency] dynamics can mask unbiological outcomes in the original model (e.g. relative abundances oscillate around equilibrium while absolute abundances shrink to zero or explode to infinity).
Indeed, the absolute abundance model used to derive our species frequency dynamics does not generally possess any fixed point, which is a basic requirement for species coexistence.
Our analysis suggests that the internal dynamics of plant or soil communities must interact with PSFs to maintain diversity in natural systems.
Multispecies coexistence becomes possible when plants, soils, or both experience an independent source of self-regulation, which might arise from resource competition, physical limits to density or some other mechanism.
Odenbaugh, 2005. “Idealized, inaccurate but successful: A pragmatic approach to evaluating models in theoretical ecology”. https://doi.org/10.1007/s10539-004-0478-6
Bever et al. 1997, “Incorporating the Soil Community into Plant Population Dynamics: The Utility of the Feedback Approach” https://www.jstor.org/stable/2960528